
ათწლეულების განმავლობაში ონკოლოგიური კვლევები მრავალრიცხოვანი, ფრაგმენტული გენეტიკური მუტაციებისა და ჰეტეროგენული უჯრედული ანომალიების რთულ ერთობლიობას წარმოადგენდა. მეცნიერები ამ ვრცელ მონაცემთა ბაზებში ფუნდამენტური კანონზომიერების პოვნასა და პათოლოგიური პროცესის ერთიანი იერარქიის ჩამოყალიბებას ცდილობდნენ. ვითარება 2000 წელს პრინციპულად შეიცვალა, როდესაც დუგლას ჰანაჰანისა და რობერტ ვაინბერგის მიერ გამოქვეყნებულმა ნაშრომმა მანამდე არსებული ინფორმაციული დისპერსია მკაფიო კონცეპტუალურ პრინციპებად დააჯგუფა.
ავტორებმა ექვსი „დამახასიათებელი ნიშნის“ (hallmarks) თეორია წარადგინეს, რაც იმ შეძენილ ფუნქციურ შესაძლებლობებს აღწერდა, რომლებიც სიმსივნეს პროლიფერაციის, ინვაზიისა და თერაპიული რეზისტენტობის მოპოვების საშუალებას აძლევდა. ამ სისტემურმა ხედვამ თვალსაჩინო გახადა, თუ როგორ აღწევს სიმსივნე ორგანიზმის ჰომეოსტაზურ მექანიზმებზე სრულ კონტროლს და რა მრავალსაფეხურიან ევოლუციურ გზას გადის იგი ბიოლოგიური ავტონომიურობის მოსაპოვებლად.
ოცდაექვსი წლის შემდეგ ეს ნაშრომი კიდევ უფრო გამდიდრდა და სამეცნიერო თვალსაზრისით გაძლიერდა. თანამედროვე მულტიგანზომილებიანი მოდელი ცხრა ძირითად მახასიათებელთან ერთად, ხუთ ხელშემწყობ ფაქტორს, სიმსივნის მიკროგარემოს (TME) კომპლექსურ სტრუქტურასა და მთელ ორგანიზმთან სისტემურ ურთიერთქმედებებს აერთიანებს. მსგავს კონცეპტუალურ ევოლუციას პრინციპული მნიშვნელობა ენიჭება, ვინაიდან კიბო გლობალურად სიკვდილიანობის ერთ-ერთ უმთავრეს მიზეზად რჩება, ხოლო მკურნალობის ეფექტურობა ხშირად ადაპტაციური რეზისტენტობის გამო იკლებს.
განახლებული მოდელი სიმსივნის სხვადასხვა ფორმისთვის დამახასიათებელ უნივერსალურ პათოფიზიოლოგიურ მექანიზმებს წარმოაჩენს, რაც მედიცინას მრავალმიზნობრივი კომბინირებული თერაპიების დანერგვისკენ უბიძგებს.
საწყისი ექვსი მახასიათებელი
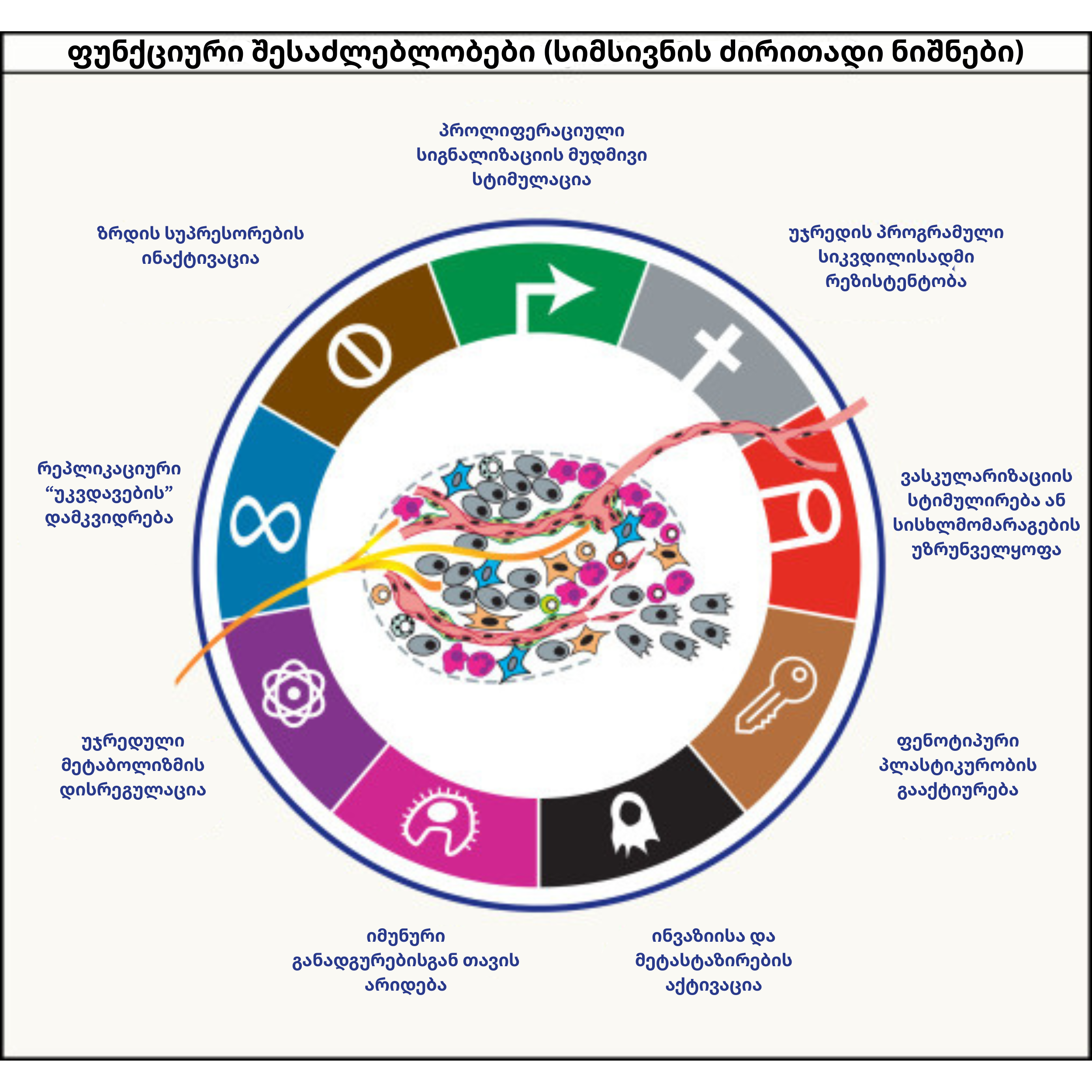
ჰანაჰანისა და ვაინბერგის მიერ იდენტიფიცირებული ექვსი თვისება იმ ბიოლოგიურ სტრატეგიებს აერთიანებს, რომელთა მეშვეობითაც სიმსივნური – ნორმალურისგან პრინციპულად განსხვავდება. ეს ნიშნები სიმსივნის ფორმირების პროცესში ეტაპობრივად ჩნდება, როდესაც უჯრედი მუტაციებს “აგროვებს” და გარემოსთან ადაპტირებს.
ეს ევოლუციური პროცესი, პირველ რიგში, ზრდის კონტროლის დაკარგვით იწყება. თუ ნორმალური უჯრედი გარეგან სტიმულს ემორჩილება, სიმსივნური უჯრედი პროლიფერაციულ სიგნალიზაციას მუდმივად, თვითონვე ინარჩუნებს. ამ ავტონომიურობას ხშირად KRAS-ისა (შემთხვევათა 30%-ში) და MYC-ის (შემთხვევათა 40%-ში) მსგავსი ონკოგენების გააქტიურება განაპირობებს, რაც სიმსივნეს დაუყოვნებელი გამრავლების შესაძლებლობას აძლევს. თუმცა, მხოლოდ ზრდის სტიმულირება საკმარისი არ არის; პარალელურად, სიმსივნემ ის ბუნებრივი „მუხრუჭებიც“ უნდა გამორთოს, რომლებიც უჯრედულ ციკლს არეგულირებენ. ამ ფუნქციას ზრდის სუპრესორების, მათ შორის TP53-ისა და RB გენების ინაქტივაცია ასრულებს, რაც უჯრედს შეკავების გარეშე გაყოფის საშუალებას აძლევს.
როდესაც უჯრედი ასეთ პათოლოგიურ მდგომარეობას აღწევს, ორგანიზმი მის განადგურებას პროგრამირებული სიკვდილის, ანუ აპოპტოზის მეშვეობით ცდილობს. სიმსივნის გადარჩენისთვის კრიტიკულია ამ მექანიზმისადმი რეზისტენტობის მოპოვება, რასაც იგი ანტიაპოპტოზური ცილების (მაგალითად, BCL-2-ის) დონის გაზრდითა და TP53-ის დამცავი ფუნქციის ბლოკირებით ახორციელებს. ამგვარად, უჯრედი არა მხოლოდ მუდმივად იყოფა, არამედ სიკვდილისგან თავდაცვის უნარსაც იძენს.
ამ პროცესს თან სდევს რეპლიკაციური პოტენციალის ზრდაც; ნორმალურ უჯრედებს ტელომერების დამოკლების გამო გაყოფის მკაფიო ლიმიტი აქვთ, რის შემდეგაც ისინი სენესცენციის (დაბერების) ან კრიზისის ფაზაში შედიან. სიმსივნეების უმეტესობა ამ ბარიერს ტელომერაზას რეაქტივაციითა და TERT პრომოტორის მუტაციით ლახავს. სხვები კი ტელომერების დაგრძელების ალტერნატიულ გზას (ALT), გენეტიკური რეკომბინაციის მეშვეობით ირჩევენ.
სიმსივნური მასის ზრდასთან ერთად, უჯრედებს ჟანგბადისა და ნუტრიენტების დეფიციტი ექმნებათ. ამ კრიზისს სიმსივნე ანგიოგენეზის სტიმულირებით პასუხობს – იგი VEGFA-სა და ANGPT2-ის სეკრეციით აიძულებს ორგანიზმს, შექმნას ახალი სისხლძარღვები, რომლებიც სიმსივნურ კერას პირდაპირ მოამარაგებენ.
საბოლოოდ კი, პროცესი ყველაზე აგრესიულ ფაზაში – ინვაზიასა და მეტასტაზირებაში გადადის. უჯრედები ქსოვილოვან ბარიერებს არღვევენ, სისხლის მიმოქცევის სისტემაში აღწევენ და დისტანციურ ორგანოებში კოლონიზდებიან. მიუხედავად იმისა, რომ ერთი კონკრეტული „მეტასტაზის გენი“ არ არსებობს, TP53 მუტაციები და ქრომოსომული არასტაბილურობა ამ პროცესს მნიშვნელოვნად აჩქარებს. ამასვე უწყობს ხელს პრემეტასტაზური ნიშები, რომლებიც დისტანციურ ქსოვილებს სიმსივნური უჯრედების მისაღებად წინასწარ “ამზადებენ”.
მეტაბოლური რეპროგრამირება და იმუნური ტოლერანტობა
2011 წლის განახლებაში ჰანაჰანმა და ვაინბერგმა კიბოს ბიოლოგიის კიდევ უფრო ღრმა შრეები წარმოაჩინეს. ტექნოლოგიურმა პროგრესმა მეცნიერებს საშუალება მისცა, დაედგინათ, რომ სიმსივნური უჯრედი არა მხოლოდ დამოუკიდებლად იზრდება, არამედ საკუთარ ენერგეტიკულ ბალანსსაც რადიკალურად ცვლის.
ამ კონტექსტში მეშვიდე მახასიათებელი უჯრედული მეტაბოლიზმის დისრეგულაციას გულისხმობს. ნორმალური უჯრედები ენერგიას ეფექტურად, ჟანგვითი ფოსფორილირების გზით გამოიმუშავებენ, პროლიფერაციული სიმსივნური უჯრედები კი ჰიბრიდულ სტრატეგიებს ირჩევენ. ისინი გლუკოზას აერობული გლიკოლიზის, ე.წ. „ვარბურგის ეფექტის“ მეშვეობით მოიხმარენ, რათა ენერგია და ბიოსინთეზური რესურსები სწრაფი ზრდისთვის გამოიყენონ. ამავდროულად, უჯრედები სიმსივნის მიკროგარემოდან (TME) გლუტამინისა და ლაქტატის იმპორტირებას მეტაბოლური სიმბიოზის პრინციპით ახორციელებენ. ნუტრიენტული სტრესის დროს კი, კომპონენტების რეციკლირების მიზნით, ისინი აუტოფაგიის მექანიზმს მიმართავენ. მსგავს მეტაბოლურ პლასტიკურობას ონკოგენები და მიკროგარემოდან მიღებული სიგნალები (მათ შორის ჰიპოქსია) განაპირობებს.
მერვე მახასიათებელი კიბოს მიერ იმუნური განადგურებისგან თავის არიდების უნარია. ფიზიოლოგიურ პირობებში, T-უჯრედები სიმსივნის სელექციურ ელიმინაციას ნეოანტიგენების იდენტიფიცირებით ახორციელებენ, რასაც NK-უჯრედების მიერ დაბალი MHC-ექსპრესიის მქონე სამიზნეების ციტოტოქსიკური განადგურება ემატება. მიუხედავად ამისა, ანტიგენის ხანგრძლივი და უწყვეტი ზემოქმედებით სიმსივნე T-უჯრედების ფუნქციურ დაქვეითებას, ანუ გამოფიტვას განაპირობებს. პარალელურად, სიმსივნის მიკროგარემო იმუნურ პასუხს სპეციფიკური ციტოკინების, რეგულატორული T-უჯრედებისა და მიელოიდური სუპრესორების მეშვეობით თრგუნავს. ამ მექანიზმის მნიშვნელობა იმუნური გამშვები პუნქტების ინჰიბიტორებმა (anti-PD1/PD-L1, anti-CTLA4) დაადასტურეს, რომლებმაც მელანომისა და ფილტვის კიბოს მკურნალობაში ნამდვილი რევოლუცია მოახდინეს.
ამავე პერიოდში კონცეფციას ორი ხელშემწყობი მახასიათებელიც დაემატა. პირველი მათგანი გენომური არასტაბილურობის ზრდას გულისხმობს, რაც მუტაციებისა და ქრომოსომული ანომალიების დაგროვებით ახალი თვისებების შეძენის საფუძველს წარმოადგენს. მეორე ფაქტორი კი სიმსივნის ხელშემწყობი ანთებაა, რომელიც კიბოს პროგრესირებისთვის საჭირო ხელსაყრელ ბიოლოგიურ პირობებს ქმნის.
მეცხრე მახასიათებელი: ფენოტიპური პლასტიკურობა
2022 წელს, ერთუჯრედიანი ანალიზის (single-cell profiling) მეთოდებმა შიდასიმსივნური ჰეტეროგენულობის ისეთი მასშტაბები გამოავლინა, რომლებიც მხოლოდ გენეტიკური მუტაციებით არ აიხსნება. ამ აღმოჩენამ საფუძველი დაუდო ფენოტიპური პლასტიკურობის, როგორც მეცხრე დამოუკიდებელი მახასიათებლის, აღიარებას. თავის მხრივ, ეს მეცხრე ნიშანი სიმსივნური უჯრედების მიერ საკუთარი ბიოლოგიური მდგომარეობის დინამიკურ ცვლილებას გულისხმობს.
ეს პროცესი ორ ძირითად მიმართულებად იყოფა. “შიდასაგვარეულო“ (intra-lineage) პლასტიკურობის დროს უჯრედები დე-დიფერენცირებას განიცდიან და საწყის, პროგენიტორულ ან სიმსივნის ღეროვანი უჯრედების (CSC) მდგომარეობას უბრუნდებიან. პარალელურად, არსებობს ტრანს-საგვარეულო პლასტიკურობა, რომელიც ეპითელურ-მეზენქიმური ტრანზიციის (EMT) ჰიბრიდულ ფორმებს მოიცავს. მსგავსი გარდაქმნები უჯრედებს ინვაზიის უნარს მატებს და წამლებისადმი რეზისტენტობას საგრძნობლად აძლიერებს.
ამგვარ სტრუქტურულ ძვრებს გენომში TP53-ის ფუნქციის დაკარგვა უწყობს ხელს, ხოლო უშუალოდ პროცესს ეპიგენეტიკური ცვლილებები და ტრანსკრიფციული ფაქტორები წარმართავენ. აღსანიშნავია, რომ მსგავს პლასტიკურობას მხოლოდ სიმსივნური ერთეულები არ ავლენენ; მასში მიკროგარემოს სხვა კომპონენტები, მათ შორის სიმსივნესთან ასოცირებული მაკროფაგები (TAMs), აქტიურად მონაწილეობენ. სწორედ ეს ბიოლოგიური მოქნილობა წარმოადგენს იმ ფუნდამენტურ სირთულეს, რომელიც თერაპიულ რეზისტენტობას კიბოს ყველა ტიპში უდევს საფუძვლად.
ოთხგანზომილებიანი ჩარჩო
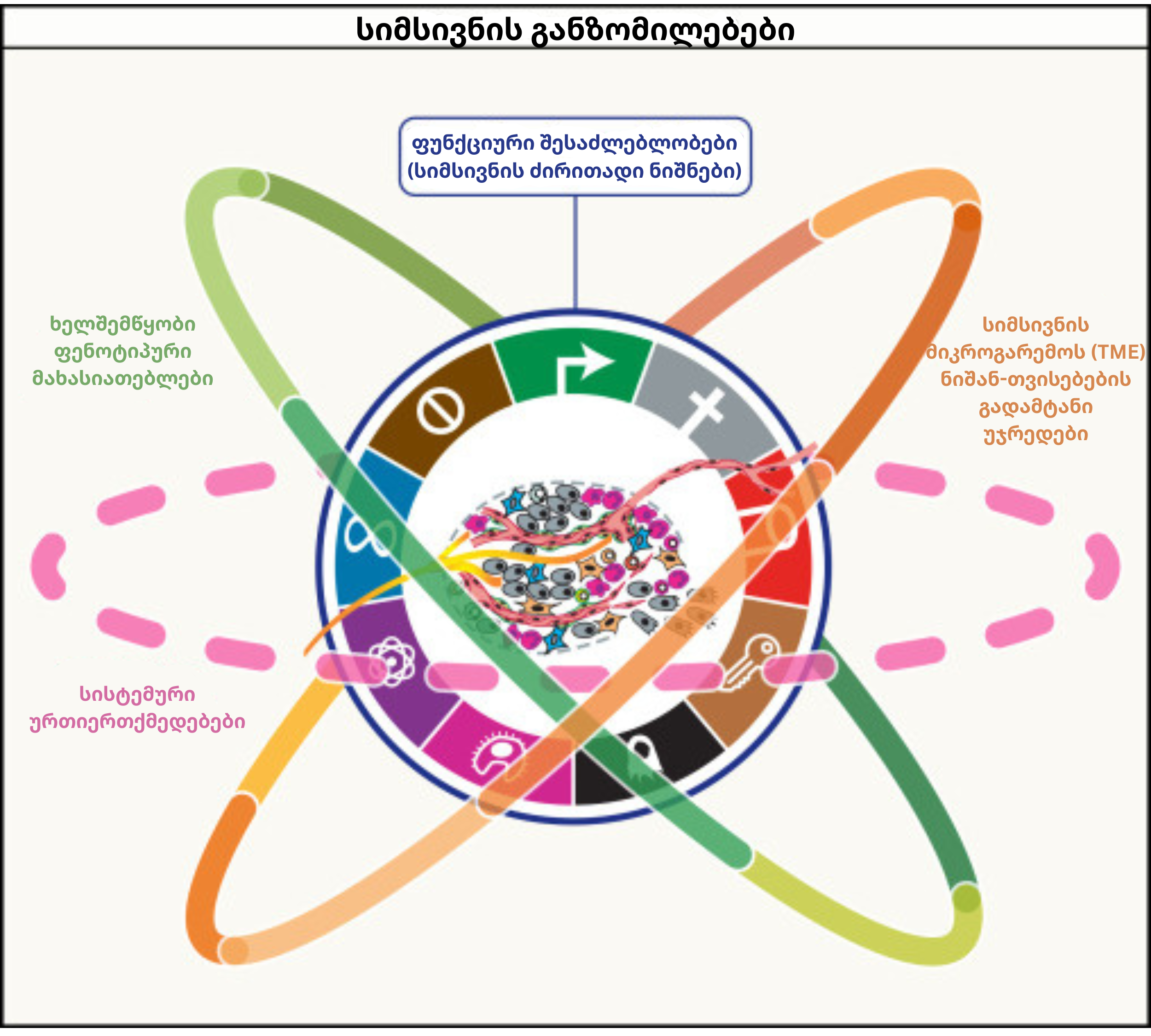
მზარდი ინფორმაციული ნაკადის პირობებში კონცეპტუალური სიცხადის შესანარჩუნებლად, თანამედროვე სამეცნიერო ხედვა დაგროვილ ცოდნას ოთხ იერარქიულ განზომილებად აჯგუფებს. ეს სტრუქტურა სიმსივნის ბიოლოგიურ სირთულეს მრავალშრიან კონტექსტში წარმოაჩენს:
პირველი განზომილება: ცხრა ფუნდამენტურ დამახასიათებელ ნიშანს (hallmarks) აერთიანებს.
მეორე განზომილება: ხუთ ხელშემწყობ მახასიათებელს მოიცავს. აღნიშნული ფაქტორები სიმსივნეს იმ გენეტიკური და ეპიგენეტიკური ცვლილებების მოპოვებაში ეხმარება, რომლებიც ზემოხსენებული ფუნდამენტური ნიშნების ჩამოყალიბებას განაპირობებს.
მესამე განზომილება: სიმსივნის მიკროგარემოს (TME) დინამიკურ შემადგენლობას აღწერს. აქ აქცენტი იმ არა-სიმსივნურ უჯრედებსა და სტრუქტურებზე კეთდება, რომლებიც პათოლოგიური პროცესების პრაქტიკულ რეალიზებას ემსახურებიან.
მეოთხე განზომილება: იერარქიის უმაღლეს საფეხურზე მთელ ორგანიზმთან მიმდინარე სისტემური ურთიერთქმედებები განიხილება. ეს განზომილება სიმსივნის გავლენას შორეულ ორგანოებზე, ნივთიერებათა ცვლასა და ორგანიზმის საერთო ჰომეოსტაზზე შეისწავლის.
ხელშემწყობი მახასიათებლები: სიმსივნური ნიშნების ფორმირების კატალიზატორები
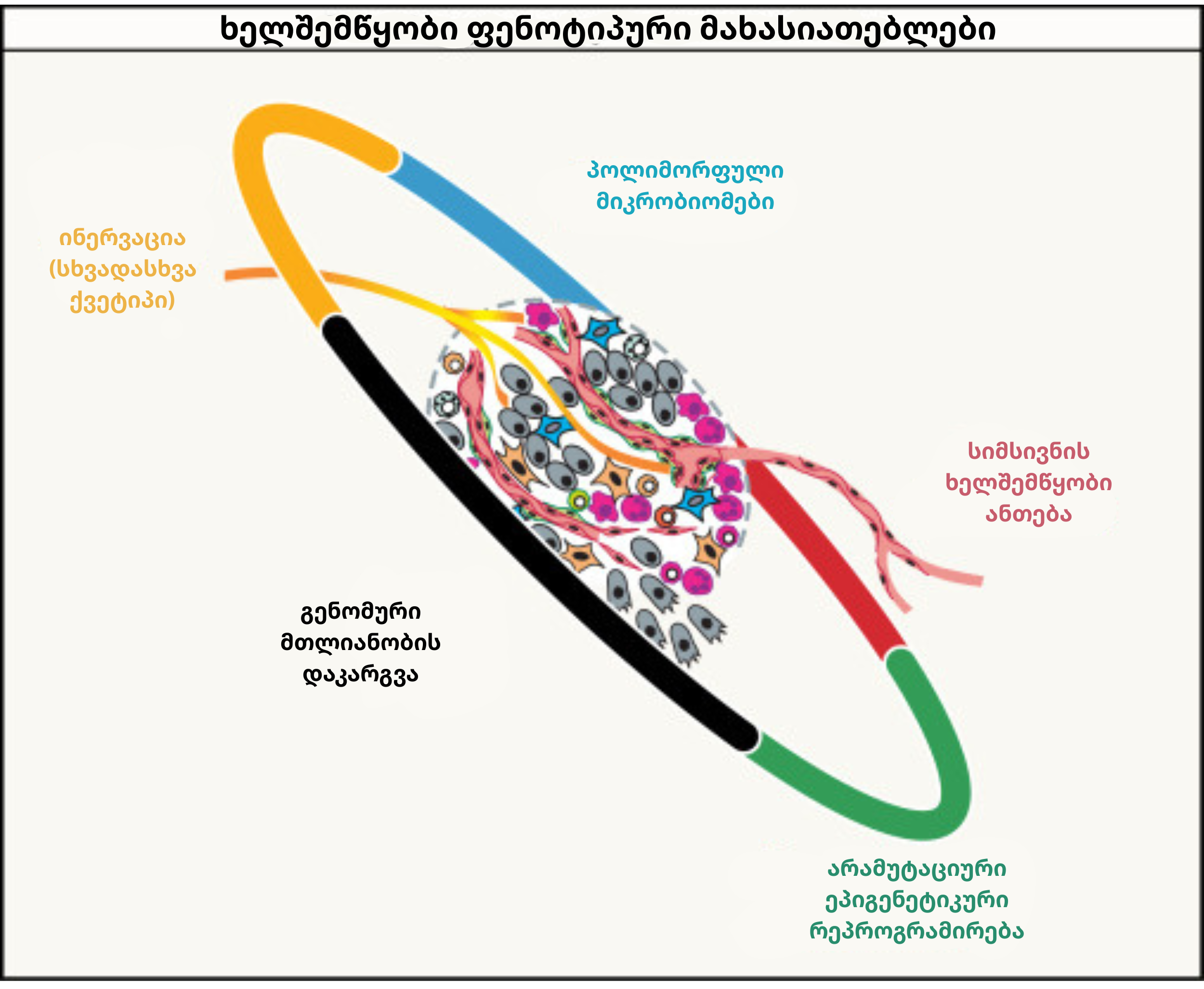
სიმსივნის პროგრესირების პროცესში არსებობს ხუთი ფუნდამენტური თვისება, რომლებიც დამახასიათებელი ნიშნების შეძენასა და მათ მდგრადობას მნიშვნელოვნად ამარტივებს:
გენომური მთლიანობის დაკარგვა: წერტილოვანი მუტაციები, ამპლიფიკაციები, დელეციები და ექსტრაქრომოსომული დნმ (ecDNA) უჯრედში გენეტიკურ ქაოსს წარმოქმნის. ამ პროცესს ქრომოტრიფსისი (ქრომოსომების ერთდროული მსხვრევა) და გენომის სრული გაორმაგებაც აძლიერებს. COSMIC-ის მონაცემთა ბაზის მიხედვით, დღეისთვის 750-ზე მეტი გენი ონკოგენად ან სუპრესორად კლასიფიცირდება. აღსანიშნავია ტელომერული კრიზისის როლიც, რომელიც გენეტიკურ მრავალფეროვნებას მანამ ზრდის, სანამ “უკვდავების მექანიზმი” მას ფიქსირებულ სტაბილურობას შესძენს.
არამუტაციური ეპიგენეტიკური რეპროგრამირება: დნმ-ის პირდაპირი სტრუქტურული ცვლილების გარეშე, ჰისტონების მოდიფიკაცია და მეთილირება გენების ექსპრესიას ფუნდამენტურად ცვლის. ეს მექანიზმი ფენოტიპურ პლასტიკურობას წარმართავს და სათავეს უდებს ზოგიერთ პედიატრიულ სიმსივნეს, რომლებსაც კლასიკური მამოძრავებელი მუტაციები აკლიათ.
სიმსივნის ხელშემწყობი ანთება: სიმსივნეები ხშირად „შეუხორცებელ ჭრილობებს“ ბაძავენ, სადაც მაკროფაგები და ნეიტროფილები დაზიანების სიგნალების (DAMPs) საშუალებით მობილიზდებიან. ეს ანთებითი გარემო ერთდროულად ხუთ სხვადასხვა ნიშანს, მათ შორის პროლიფერაციასა და ანგიოგენეზს, ასტიმულირებს. ამავდროულად, KRAS და TP53 მუტაციები მიელოიდური დამხმარე უჯრედების აქტიურ მობილიზებას უწყობს ხელს.
ინერვაცია: თანამედროვე კვლევები ადასტურებს, რომ ნერვული ბოჭკოები სიმსივნეში ღრმად აღწევენ და კიბოს უჯრედებთან პირდაპირ სინაფსებს აყალიბებენ. გამოყოფილი ნეიროტრანსმიტერები სიმსივნის გადარჩენას, ინვაზიასა და იმუნური სისტემისგან თავის არიდებას უწყობს ხელს. ამ კავშირების დასამტკიცებლად კიბოს ნეირომეცნიერება ოპტოგენეტიკის მეთოდს იყენებს. თავის მხრივ, სიმსივნეები აქსონების ინტენსიურ ზრდას ასტიმულირებენ და ნეირონების ფუნქციურ რეპროგრამირებას წარმართავენ.
პოლიმორფული მიკრობიომი: ნაწლავის, კანისა თუ უშუალოდ სიმსივნის შიდა მიკრობიომი ინდივიდებს შორის მკვეთრად განსხვავდება. გარკვეული ბაქტერიები იმუნური სისტემის დათრგუნვას უწყობენ ხელს, რასაც ICI-თერაპიისადმი რეზისტენტობამდე მივყავართ. ამის საპირისპიროდ, ფეკალური ტრანსპლანტაცია მკურნალობის გამოსავალს საგრძნობლად აუმჯობესებს. დისბიოზი კი, თავის მხრივ, მუტაციური პროცესებისა და ანთების ინტენსივობას კიდევ უფრო ზრდის.
სიმსივნის მიკროგარემო
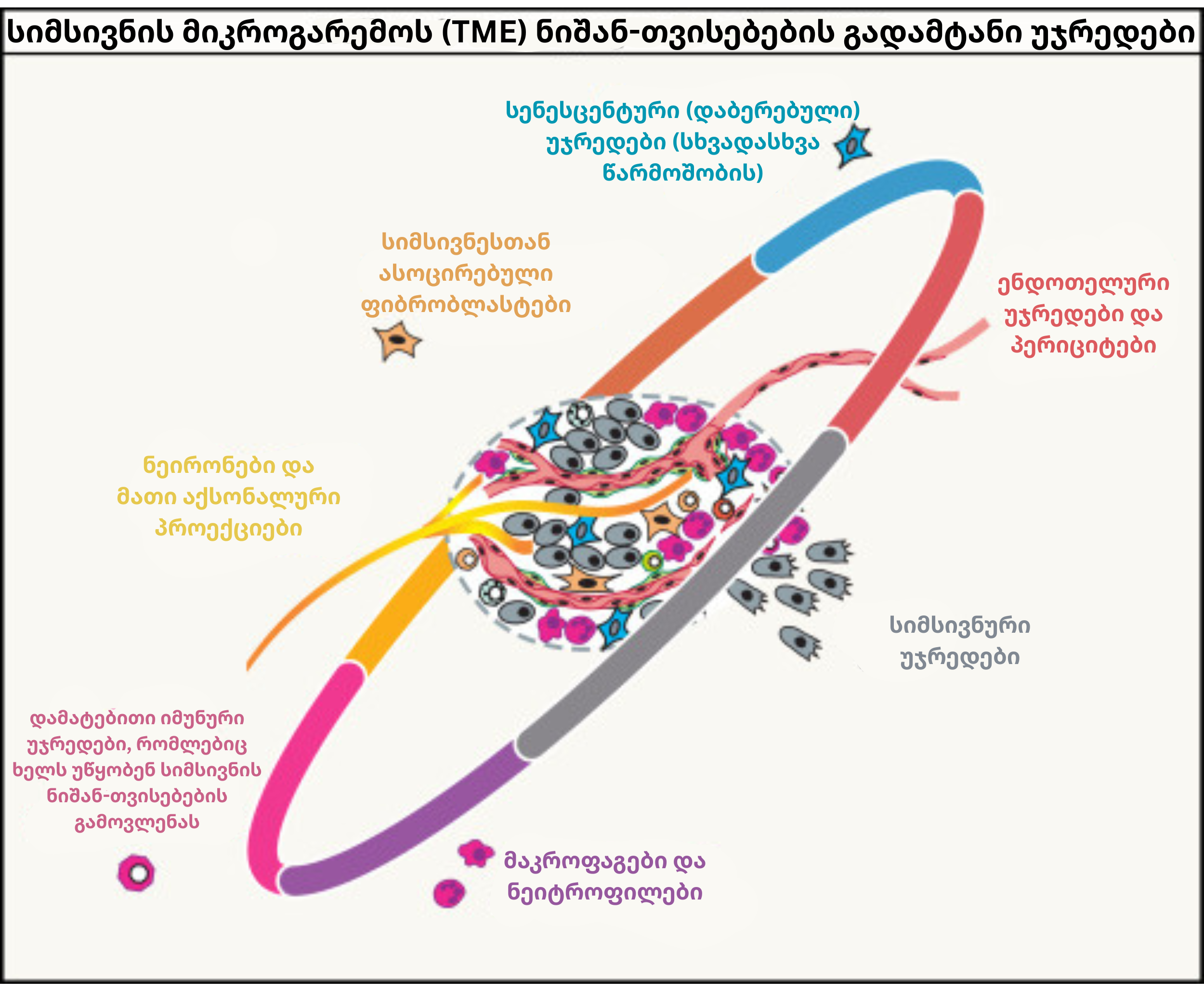
თანამედროვე ხედვით, სიმსივნე ფუნქციონირებს როგორც რთული პათოლოგიური ორგანო, რომელიც ნორმალურ უჯრედებს საკუთარი ინტერესებისთვის იყენებს. ამ გარემოში თითოეული კომპონენტი დაავადების პროგრესირებას თავისებურად ემსახურება:
სიმსივნური უჯრედები: ისინი დაავადების გენეტიკურ ბირთვს წარმოადგენენ, მამოძრავებელ მუტაციებს ატარებენ და შეიცავენ სიმსივნის ღეროვანი უჯრედების (CSC) რეზერვუარებს. ეს უკანასკნელნი თერაპიულ ჩარევას მიძინებული (dormancy) მდგომარეობით უპირისპირდებიან: სანამ ქიმიოთერაპია აქტიურად გამრავლებად უჯრედებს ანადგურებს, ეს „მძინარე“ უჯრედები დროებით წყვეტენ გაყოფას, ინარჩუნებენ სიცოცხლისუნარიანობას და მკურნალობის დასრულების შემდეგ სიმსივნის ხელახალ ზრდას განაპირობებენ.
ენდოთელური უჯრედები და პერიციტები: ისინი ქმნიან სტრუქტურულად გაუმართავ სისხლძარღვებს, რომლებიც სიმსივნეს ზრდის ფაქტორებით ამარაგებენ. ამავდროულად, ეს სისხლძარღვოვანი ქსელი T-უჯრედების ინფილტრაციას ზღუდავს და მეტასტაზურ პროცესებს საგრძნობლად ამარტივებს.
კიბოსთან ასოცირებული ფიბრობლასტები (CAFs): ისინი ლოკალური ფიბრობლასტებისგან, ენდოთელურ-მეზენქიმური ტრანზიციისა (EndMT) ან ძვლის ტვინის ღეროვანი უჯრედებისგან ყალიბდებიან. მათ შორის მიოფიბრობლასტური CAFs უჯრედგარე მატრიქსს ამკვრივებს, ხოლო ანთებითი CAFs სპეციფიკურ ციტოკინებს გამოყოფს და მეტაბოლურ აქტივობას ასტიმულირებს.
TAMs და TANs: სიმსივნესთან ასოცირებული მაკროფაგები და ნეიტროფილები მაღალი პლასტიკურობით ხასიათდებიან; ისინი სიმსივნის საწინააღმდეგო მდგომარეობიდან მის ხელშემწყობ ფაზამდე ტრანსფორმირდებიან. მაგალითად, M2-ტიპის მაკროფაგები ანგიოგენეზსა და ინვაზიას წარმართავენ, ხოლო მიელოიდური სუპრესორული უჯრედები (MDSCs) T-ლიმფოციტების ფუნქციას აქტიურად თრგუნავენ.
იმუნური უჯრედები: რეგულატორული T-უჯრედები (Tregs), დისფუნქციური დენდრიტული უჯრედები და NK უჯრედები სიმსივნის მიკროგარემოში საკუთარ პირვანდელ დანიშნულებას კარგავენ. ნაცვლად დამცველობითი რეაქციისა, ისინი იმუნოსუპრესიული გარემოს ფორმირებასა და სიმსივნის ხელშეუხებლობას უზრუნველყოფენ.
ნეირონები: ნეირონები სიმსივნურ უჯრედებთან პირდაპირ სინაფსებსა და ადჰეზიურ კავშირებს აყალიბებენ, რის შემდეგაც, ნეიროტრანსმიტერების გამოყოფით, კიბოსთვის დამახასიათებელი თვისებების გამომწვევ სიგნალებს გასცემენ.
სენესცენტური უჯრედები: ნებისმიერი წარმოშობის დაბერებული უჯრედები SASP-ფაქტორებს გამოყოფენ, რაც პარადოქსულად პროლიფერაციას, ანგიოგენეზსა და ანთებას მეზობელ ერთეულებში აღვივებს.
აღნიშნული უჯრედების დესტრუქციულ წვლილს ისეთი ფიზიკური ფაქტორებიც აძლიერებს, როგორიცაა მატრიქსის მაღალი სიმკვრივე, ჰიპოქსია და გარემოს მჟავიანობა.
სისტემური ურთიერთქმედებები – კიბო, როგორც მთელი ორგანიზმის დაავადება
ონკოლოგიური პათოლოგია მხოლოდ ლოკალური სიმსივნით არ შემოიფარგლება. თანამედროვე მედიცინა მას განიხილავს, როგორც სისტემურ პროცესს, რომელიც ორგანიზმის საერთო ბიოლოგიურ ფონთან მჭიდროდ ურთიერთქმედებს:
დაბერება და იმუნური ფონი: დაბერების პროცესი ორგანიზმში მუტაციების დაგროვებასა და იმუნიტეტის საყოველთაო დაქვეითებას იწვევს. ამავდროულად, მატულობს სენესცენტური უჯრედების რაოდენობა, რომლებიც პრო-სიმსივნურ SASP-ფაქტორებს გამოყოფენ. ეს ცვლილებები კიბოს დამახასიათებელი ნიშნების ერთგვარ ბიოლოგიურ საფუძველს წარმოადგენს და დაავადების განვითარებისთვის ხელსაყრელ ნიადაგს ამზადებს.
მეტაბოლური გავლენა და სიმსუქნე: სიმსუქნე ორგანიზმში ლეპტინის დონეს ზრდის და ადიპონექტინის სინთეზს თრგუნავს. აღნიშნული ჰორმონალური დისბალანსი პროლიფერაციას, ანგიოგენეზს, ქრონიკულ ანთებასა და მეტაბოლურ ძვრებს პირდაპირ ასტიმულირებს.
დისტანციური კომუნიკაცია: დისრეგულირებული მიკრობიომი ორგანიზმის სხვადასხვა სისტემაზე დისტანციურ გავლენას ავრცელებს, მაშინ როცა ინერვაცია და ვასკულარიზაცია სიმსივნესა და შორეულ ორგანოებს შორის ერთიან საინფორმაციო და სატრანსპორტო ქსელს ქმნის.
კლინიკური გამოვლინებები: აღნიშნული კომპლექსური ურთიერთქმედებები ხშირად კუნთოვანი მასის მკვეთრ განლევაში და იმუნური სისტემის საერთო დასუსტებაში ვლინდება. ამ დესტრუქციულ პროცესებს გარემო ტოქსინები (მათ შორის თამბაქოს კვამლი) ანთებითი რეაქციების გაძლიერებისა და პროცესების ესკალაციის გზით კიდევ უფრო ამძაფრებს.
2020-იანი წლების მიღწევები
ეს უახლესი ეტაპი იმ ტრანსფორმაციულ აღმოჩენებს აერთიანებს, რომლებმაც სიმსივნის აღქმა „გენეტიკური შეცდომიდან“ რთულ, მრავალშრიან ეკოსისტემამდე აიყვანა. 2020-იანი წლების მთავარი მიღწევა სწორედ ამ იზოლირებული ნიშნების ერთიან, ინტერაქციულ ქსელში გაერთიანება აღმოჩნდა. თუ ადრე თითოეულ მახასიათებელს (მაგალითად, ინერვაციას ან მიკრობიომს) ცალკეულ ფენომენად განვიხილავდით, დღეს მეცნიერება მათ შორის არსებულ მჭიდრო იერარქიულ კავშირებსა და ფუნქციურ გადაკვეთებს გამოყოფს. თუმცა, ყველაზე მნიშვნელოვანი მაინც დადასტურებული „კო-თარგეთინგის“ (ერთდროული მიზანმიმართული თერაპიის) სტრატეგიებია, რომლებიც თერაპიული ეფექტის შენარჩუნებისა და დაავადების ხანგრძლივი რემისიის რეალურ შესაძლებლობას ქმნის.
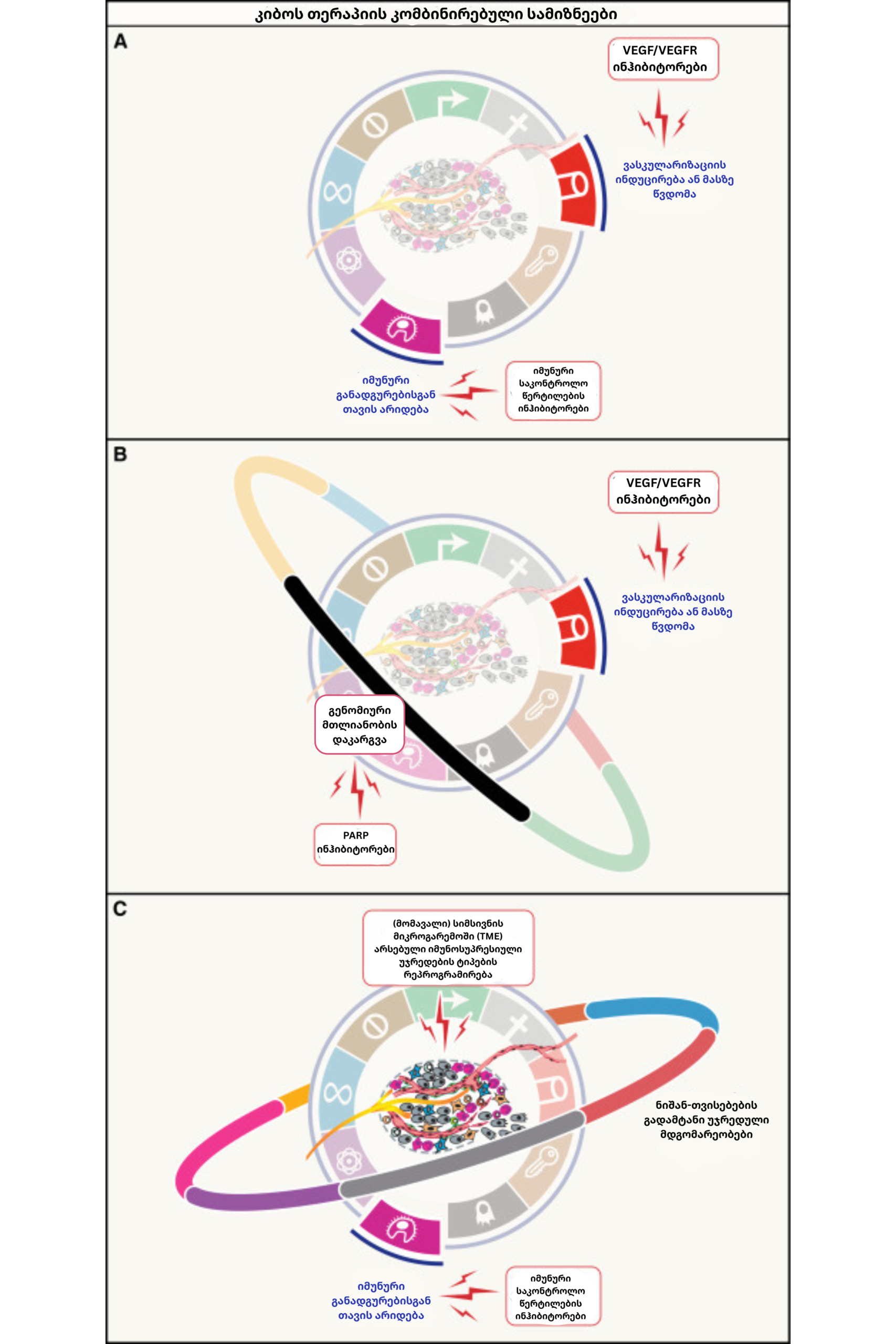
ამ ფონზე, ონკოლოგიის მომავალი მოწინავე ტექნოლოგიებისა და პერსონალიზებული მიდგომების სინთეზს ეფუძნება. დაავადების მართვა ახლა უკვე პროგნოზირებად მოდელებზე გადადის, სადაც ერთუჯრედიანი მულტი-ომიქსი, სივრცითი ტრანსკრიპტომიკა და ხელოვნურ ინტელექტზე დაფუძნებული ციფრული პათოლოგია სიმსივნის ქცევასა და მკურნალობაზე მოსალოდნელ პასუხს მაღალი სიზუსტით განსაზღვრავს. გარდა ამისა, სამომავლო თერაპიული სტრატეგიები სიმსივნის რამდენიმე დამოუკიდებელ ნიშანზე ერთდროული მიზანმიმართული ზემოქმედებისკენ იქნება მიმართული, რაც მკურნალობისადმი რეზისტენტობის ჩამოყალიბების ალბათობას მნიშვნელოვნად ამცირებს.
წყარო: cell
